
Zoonotic spillovers: How climate change, habitat destruction, and bushmeat trade might amplify bat-driven viral disease risks
- Review
- Published:
- Volume 71, article number 72 (2025)
- Cite this article
Abstract
Bats are indispensable members of the natural world, supporting its delicate balance. Bats have vital roles in controlling insect populations and enhancing soil fertility. They also help in the harvesting and dispersal of seeds, pollination in plants, and nutrient recycling and distribution. However, through evolution over millions of years, they have also adapted their immune system so that they may carry numerous types of pathogens, the majority of which are viruses, without these pathogens having any serious ill effects on bats themselves. Their anatomical adaptation to flight and the reduced immune response to DNA damage during flight have also contributed to bats becoming reservoirs of deadly pathogenic diseases. This review discusses the different adaptations of bats with a special focus on the immune system that have helped them evolve as a reservoir for various viruses. The study also enumerates how the increase in global warming, the consequent changes in climatic conditions, habitat destruction, and bushmeat consumption increase the chances of an outbreak of novel zoonotic disease when humans come in contact with bats.
Graphical abstract
Similar content being viewed by others
Bats as Reservoirs of Viral Zoonoses
{kind=link}
Viral ecology in chiroptera: human-wildlife interactions and pandemic risk
Explore related subjects
Discover the latest articles, books and news in related subjects, suggested using machine learning.Introduction
Bats can be the reservoir host of different pathogens, particularly some viruses including various species of the genus Lyssavirus, Influenzavirus A, Henipavirus (e.g. Hendra virus, Nipah virus), Flavivirus (e.g. Japanese Encephalitis Virus, St Louis Encephalitis Virus, West Nile virus, Dengue virus), Alphavirus (e.g. Venezuelan equine encephalitis virus, Chikungunya, Sindbis virus), Rhabdovirus (e.g. Australian bat lyssavirus), Coronaviruses (e.g. SARS-CoV, SARS-CoV-2, MERS-CoV), Filoviruses (e.g. Marburg virus), and Sosuga virus (Amman et al. 2014; Mackenzie et al. 2016; Liu et al. 2024). Bats have been identified to play an important role in the evolution of coronaviruses (CoV) and are known to host at least 30 different CoVs (Platto et al. 2021). According to the WHO (2022) guidelines, and Weinberg and Yovel (2022), bat-borne diseases such as Ebola, Hendra virus infection, Influenza, Marburg disease, MERS-CoV (Middle East respiratory syndrome coronavirus), Nipah virus infection, Coronavirus disease 2019 (COVID-19), and SARS (severe acute respiratory syndrome) have the potential for future epidemics or pandemics.
Interestingly, possible zoonotic transmission of these outbreaks, mostly as epidemics or pandemics, is suspected through different wildlife reservoirs, bats being one of them (Tabish and Nabil 2022). The majority of these diseases are driven by human activities such as habitat fragmentation, agricultural development, land use, and uncontrolled urbanization, which influence the transmission of infectious diseases from animals to humans (Olivero et al. 2020; Platto et al. 2020) (Fig. 1). Apart from this, hunting, wet markets, and the consumption of bat meat play a major role in transmitting bat-borne diseases (Epstein and Field 2015; Platto et al. 2020) (Fig. 1). The main reason for cross-species disease transmission stems from new and unintended interactions between various species (Galindo-González 2024). Valitutto et al. (2020) discovered three new alphacoronaviruses, three novel betacoronaviruses, and one previously identified alphacoronavirus in bats for the first time. It is highlighted that continuous changes in land use are a major contributor to the increase in zoonotic diseases, as they result in more frequent human-wildlife interactions, thus elevating the risk of zoonoses (Galindo-González 2022). Furthermore, Ye et al. (2020) investigate how interactions between CoVs and their animal hosts could offer valuable insights into CoV pathogenesis in humans. Researchers analyze various HCoVs from the perspectives of virus evolution and genome recombination, emphasizing the need to understand the conditions that facilitate successful host transitions and the influence of viral evolution on the severity of diseases.
Climate change, habitat destruction, and consumption of bushmeat, also sold in live markets, are drivers for the emergence and re-emergence of bat-related diseases
This study was undertaken with the hypothesis that bats have immune systems similar to other mammals and bat-human interaction has not increased. The following questions were considered during the study: If the bat’s immune system differs from other mammals, does it help to adapt as a carrier of microorganisms? If the bat-human interactions have increased, what are the reasons for it?
Methodology
The three most commonly used science-based databases, Google Scholar, ScienceDirect, and PubMed were used to search pertinent literature using the terms or combination of terms like “adaptation; bats; bush meat; climate change; conservation; disease reservoirs; emerging diseases; evolution; habitat destruction; immunity; interactions; pathogens; viruses; and zoonoses”. The authors rigorously filtered the original articles from the databases, websites, and organizations published till March 2024. Based on the same title, the duplicate results were quickly removed.
The relevant papers were later selected by reading the title, abstract, and keywords. For the methodical synthesis of the current study, all pertinent articles’ full texts were accessed after an initial screening. Inclusion criteria: articles covering the antiviral, host defense, and immune properties of bats; the ability of the immune system of the bats to suppress the virulence response of pathogens; anthropogenic factors responsible for increased human-bat interactions; and probable steps that can be taken to avoid unwanted disease outbreaks that may originate when an increase in human-bats interaction occurs—exclusion criteria: abstracts from seminars or conferences, viewpoints, opinions, letters, and encyclopedias. All the selected articles were further screened independently by the two authors and were discussed thoroughly for the inclusion of the relevant pieces of information. The reporting of this review is based on the directions of the systematic reviews (PRISMA) guidelines (Haddaway et al. 2022). In Fig. 2, the screening procedure is schematically illustrated. Many mammals, like rodents, pangolins, primates, bats, and pigs, can transmit zoonotic diseases to humans. However, bats are the only mammals among them which is capable of flight, found in a large area of the globe, and have vital roles in the ecosystem, making them one of the most important mammals capable of acting as a reservoir for microbes. Bats exhibit several critical characteristics that facilitate the accumulation of pathogenic organisms: (1) They are highly social animals, often forming colonies that can consist of hundreds of thousands or even millions of individuals; (2) they have a relatively long lifespan, typically ranging from 15 to 20 years or more; (3) they possess the ability to fly, with some species exhibiting migratory behavior; (4) they have a highly developed immune system (Li et al. 2005; Calisher et al. 2006; Han et al. 2015). Thus, the study was focused first on finding the specific characteristics of the bat’s behavior, anatomical adaptations, and immune system adaptations that might help suppress the effect of the micro-organisms’ virulence in their body. This was followed by searching for the anthropogenic factors that increase the chances of frequent human-bat contact and might increase the probability of disease outbreaks. To concisely present the information in an understandable and simpler form, some examples included in this study have been explained through diagrammatic and data representations of the Nipah and rabies virus epidemiology.
PRISMA flow diagram of the citation search and selection strategy. Adopted from Haddaway et al. 2022
Bats as reservoirs of pathogens
Many wild animals have the natural tendency to harbor different types of pathogens, bats are just one among them. In population, bats are the second-most abundant group of mammals after rodents. About 1462 living bats comprise approximately 22% of all known living mammal species (Burgin et al. 2018; Simmons and Cirranello 2022). The diversity of bat species (species richness) can be favorable for the emergence of new viruses that infect humans and other domestic and wild mammals (Cohn 2020; Mollentze and Streicker 2020). Compared to other mammals, bat’s unique physiological and life-history traits, such as long life span (Wilkinson and South 2002), flight ability and echolocation (Norberg and Rayner 1987), hibernation, geographical distribution, and daily torpor could influence the noticeable high viral richness for bats (Calisher et al. 2006). These factors may underlie or contribute to the immune mechanisms that help bats mount their hardy innate immune response against pathogens.
Some bat species also undergo hibernation. For such bats, emerging from hibernation is a difficult experience (Lee and McCracken 2002). Gamma herpes viruses are latently present in many big brown bats (Subudhi et al. 2018). According to Gerow et al. (2019), when large brown bats emerge from hibernation, the virus reactivates from latency, which results in the virus being found in the blood. Low levels of antiviral antibodies were also linked to this reactivation. However, after some time of the bats’ hibernation, the virus again enters latency due to rising antibody levels. Eventually, antiviral immunity increases by reducing viral DNA in the blood cells of non-hibernating bats (Gerow et al. 2019).
The sudden resurgence of some specific pathogens of bat origin, like the Nipah virus infection, is suspected to be triggered by a combination of factors such as reduced antibody levels and increased stress levels due to habitat destruction (Sohayati et al. 2011). Plowright et al. (2016) introduced a hypothesis stating that viruses infect vulnerable, naïve bats, resulting in acute illness. This then develops into a latent or chronic infection. After that, the virus periodically reactivates in response to various environmental and physiological cues. This hypothesis is known as the Susceptible–Infectious–Latent–Infectious (SILI) hypothesis (Plowright et al. 2016). The immune system of bats is unique and viral infection is likely to occur in many bats (Fig. 3).
Effect of stresses on chronic viral infection. Viral shedding in bats occurs as stressful events change the balance between host and virus and accelerate viral replication. Viruses also infect bats intently because of their unique antiviral immune response and their suppression of severe pathological effects
The antiviral and immune defense response in bats
Bats are an ancient species, dating back 64 million years. The long evolutionary history has contributed to various viruses co-evolving with bats to become their natural reservoir (Teeling et al. 2005). When challenged with the virus, Jamaican fruit bats (Artibeus jamaicensis) show no or minimal signs of disease, even when high viral loads are detected in tissues (Munster et al. 2016). Interferons (IFNs) are cytokines that induce an antiviral state in host cells and are especially responsible for resisting viral infection. Bats exhibit a distinctive IFN, which may illustrate their efficiency in coexisting with pathogens (Ahn et al. 2016). Generally, mammals carry a large IFN locus comprising a family of IFN-α genes expressing themselves only during a viral infection. In contrast, bats have only three IFN-α genes continuously and constitutively expressed (Ahn et al. 2016). In fact, except for pigs, no other mammal has high expression levels of the IFN-ω subfamily (Schountz et al. 2017; Jennings and Sang 2019). In mice, IFN-ω is lacking. When compared with other mammals, IFNω and IFNδ genes appear expanded in the black flying fox (Baker and Zhou 2015). According to the hypothesis of Pavlovich et al. (2020), the IFN-ω subfamily’s expansion might help to create a more adaptable antiviral response that would benefit bats by preventing excessive pathology. Dysregulation of the IFN response has already been involved in autoimmune diseases and the pathogenesis of various bat-borne viruses, including Ebola virus, SARS-CoV (Liu et al. 2017), and SARS-CoV-2 (Huang et al. 2020). Not all bats constitute IFN all the time (Weinberg and Yovel 2022). Conceivably, bats can manage a distinctive balance between the levels of the two key type I IFNs, assisting in their antiviral abilities (Clayton and Munir 2020). In bats, this may be managed by IRFs (IFN regulatory factors) as differential expression patterns of IRF7 (Zhou et al. 2014) and IRF3-mediated enhanced antiviral responses (Banerjee et al. 2020). This confined induction of type I IFNs would reduce the production of inflammatory cytokines (Hölzer et al. 2019). The kinetics of the IFN response in bats is also different from that of other mammals, with a rapid decline phase for some bat ISGs (Interferon-stimulated genes) (La Cruz-Rivera et al. 2018). Further, various antiviral genes, for example, RNase-L (2–5 A-dependent endoribonuclease) genes, are IFN-inducible in bats (La Cruz-Rivera et al. 2018) but not in other mammals (Hölzer et al. 2019). Additionally, when bats are stimulated by virus infection, they maintain a high basal rate of autophagy that acts as both an anti-viral and pro-survival mechanism (Laing et al. 2019). Brook and Dobson (2015) suggested that bats evolved a secondary anti-viral role for the apoptotic removal of damaged mitochondrial DNA, resulting from increased oxidative stress during flight. Simultaneously, these bat-specific changes in baseline expression, dynamics, functions, or induction of antiviral genes in IFN signaling may help bats effectively control the abundant viruses they host (Irving et al. 2021).
In addition to the innate immune responses, some bats express very high levels of heat-shock proteins (HSPs), which enable bat cells to survive high temperatures and high oxidative stress in vitro (Phillips et al. 2017). By tolerating some viral mutations and modulating viral proteins, HSPs contribute to the rapid acceleration of viral evolution (Phillips et al. 2017). It also acts as a viral receptor (Reyes-del Valle et al. 2005), regulates or blocks apoptosis (Beere et al. 2000), inflammation (Srivastava 2002), and affects aging (Singh et al. 2006). A recent study has demonstrated that bats express ABCB1 (ATP binding cassette subfamily B member 1 gene) comparatively more than humans (Koh et al. 2019). The broad expression of the ABCB1 gene uniquely promotes resistance to DNA damage induced by the chemotherapeutic drugs doxorubicin and etoposide, providing resistance to genotoxic compounds, regulating cellular homeostasis, and probably lowering the occurrence of cancer (Koh et al. 2019). Another feature is their close association with viruses (Barton et al. 2007). The pool of viruses present in the guts of bats can help the microbiome boost immunity (Kapp et al. 2010). This has also led to the survival of diverse pools of CoV quasi-species in bat populations (Seronello et al. 2011).
Unique immune properties and suppression of strong pathological effects of virus-induced inflammation
Although bats are well-equipped to control viral infection, they also have mechanisms to reduce the induction of inflammatory genes (Subudhi et al. 2019). Excessive inflammation can be harmful and is associated with unwanted pathological changes in other species of vertebrates, such as humans (Channappanavar and Perlman 2017). Bats have evolved mechanisms for controlling excessive inflammation. Studies on cell culture have revealed that the promoter region of tumor necrosis factor-alpha (TNFα), a crucial inflammatory cytokine, contains a binding site for the inhibitor molecule (cRel) in the cells of many species of bats. In Eptesicus fuscus (Big brown bat) cells, a synthetic double-stranded viral RNA (dsRNA), poly I: C (Polyinosinic: polycytidylic acid), stimulated with cRel actively suppressed TNFα expression (Banerjee et al. 2017). Genome analysis of David’s myotis and black flying foxes has also exhibited the presence of positive selection pressure on the cRel gene (Zhang et al. 2013). This indicates that many bats can maintain a balanced response to viral infection by suppressing the expression of TNFα (Subudhi et al. 2019).
STING or stimulator of interferon genes, the most adapted protein, senses damaged DNA or dsRNA from viruses and enhances an interferon response (Xie et al. 2018). Several bat species showed immune tolerance by reducing STING-dependent type I IFN activation. This adaptation has been caused by replacing the functionally important and extremely conserved serine residue at position 358 (S358) (Xie et al. 2018). The replacement of S358 by other amino acids in bat STING dampened but did not completely reduce the function of STING (Mandl et al. 2018). A weakened but not completely lost function of STING may have profound implications for bats in maintaining a balanced state of “effective response” but not “over-response” against viruses (Xie et al. 2018). The PHYIN (Pyrin and HIN domain) genes, responsible for the formation of the inflammasome and microbial DNA sensing, are not present in bats (Ahn et al. 2016). The complete loss of PYHIN in bats suggests a vital adaptation for flight (Dempsey and Bowie 2015). Therefore, its removal may enable some bats to limit excessive inflammation activation and regulate the type I IFN response to normally recognized DNA damage by PYHIN proteins. There is the possibility that the increased exposure of bats to many zoonotic RNA and DNA viruses compared to other mammals (that do not cover large distances), may have been an evolutionary driver of PYHIN loss or, conversely, that PYHIN loss may have allowed for this bat-viral co-evolution (Ahn et al. 2016). It is hypothesized that the reason for bat longevity is the loss of the PYHIN gene (Clayton and Munir 2020). These regulations suggest that bats may experience some reduction in inflammation at the time of viral infection (Subudhi et al. 2019). For the mitigation of the deleterious effects of flight, mechanisms to regulate inflammation may have evolved. Cytosolic DNA is abundantly produced in bat cells during flight, which may create a strong natural selection pressure to reduce the activation of bat DNA sensors (Subudhi et al. 2019). Similarly, a study discovered a suppressed NLRP3 (NLR-family pyrin domain-containing 3) inflammasome activity in the immune cells of MERS-CoV-infected black flying fox, which can recognize various cellular stresses and pathogen invasions and results in lower production of the proinflammatory cytokine IL-1β (Interleukin-1β) (Ahn et al. 2019). Ahn et al. (2019) showed an overall dampened activation of NLRP3 in bat immune cells compared to humans in their study (Fig. 4). Consequently, reducing immunopathology during viral infections and diminished STING and NLRP3 inflammasome activity in black flying foxes potentially evolved to decrease the pathological consequences of oxidative DNA damage during flight (Baid et al. 2024). Flights increase the metabolic rate and result in the formation of higher levels of oxygen-free radicals (Maina 2000). This makes bats more susceptible to producing damaged DNA (Cadet and Wagner 2013). Since mounting an immune response is energetically costly (Sheldon and Verhulst 1996) and harmful, bats have likely evolved mechanisms to suppress the activation of the immune system caused by damaged DNA generated during flight, thereby reducing the overall inflammation (Jebb et al. 2018). During hibernation, bats conserve energy by lowering their body temperature and metabolic rate, which can suppress the immune system and delay viral clearance (George et al. 2011).
The comparison between bat and human inflammatory responses. Bats maintain an evolutionary adaptation whereby the sensing of cytoplasmic DNA is dampened by the loss of a DNA sensor belonging to the PYHIN gene family, leading to the dampened NLRP3 inflammatory response. This loss also affects the function of STING, in which the serine residue is mutated in bats and thus has reduced function. All of these sequences appear to be unique to bats, and all contribute to the low levels of IFN production in bat cells
Major potential causes for disease outbreaks from bats
Global warming and climate change
Climate change may contribute to the risk of pathogen spillover. Environmental changes can alter species ranges and densities, leading to novel interactions between species and increasing the risk of zoonotic introduction (Carlson et al. 2022; Guth et al. 2020; Galindo-González 2024). The El Niño-Southern Oscillation (ENSO) is an important source of annual global climate variability, characterized by three phases: El Niño (warming), La Niña (cooling), and the Neutral phase (Hanley et al. 2003). ENSO is one of the most important climate events due to the global atmospheric circulation and its ability to change global temperature and precipitation patterns (Hanley et al. 2003). It directly supports outbreaks or the spread of various public health concerns (Anyamba et al. 2019). According to the WHO (2023a), El Niño occurrences have increased in South and Southeast Asia due to less rainfall. ENSO-related drought conditions and anthropogenic forest fires may have resulted in fruit bat infection with the Nipah virus while they were migrating from Sumatra to Malaysia in mid-1997 in search of food (Chua et al. 2002). The populations of black flying foxes in Australia, a main reservoir of the Hendra virus, have shifted 100 km south over the past 100 years due to climate change. This shifting range likely contributed to the spread of the Hendra virus in southern horse populations, and these horses subsequently infected humans (Yuen et al. 2021). Latinne and Morand (2022) showed in their experiment that the cool phase or La Niña event directly influenced the five emerging outbreaks including, the Nipah virus in Malaysia and India, the Melaka virus in Malaysia, SADS-CoV (Swine acute diarrhea syndrome coronavirus) in China, and MERS-CoV in Saudi Arabia, while four of them, i.e., Hendra virus in Australia, SARS-CoV-1 and − 2 in China, Menangle virus in Australia occurred at the time of warm phase or El Niño event (Table 1). The other three bat-related events, including the Australian bat lyssavirus in Australia, the Nipah virus in the Philippines, and the Kampur virus in Malaysia, occurred during the neutral phase (Table 1). Fruit bats could respond to severe weather events and climate change by shifting their distribution to more favorable areas since they are highly proactive and often travel great distances in search of resources (Roberts et al. 2012). Bats can colonize previously uninhabited areas (Boardman et al. 2020). It is predicted that in Australia, fruit bats may migrate as far south as Tasmania in the next 10 to 20 years due to the climate crisis, possibly leading to human-wildlife conflict (Diengdoh et al. 2022).
In the Americas, bat-borne rabies virus spillover from wildlife to domesticated species is substantial (Pan American Health Organization 2024). In Latin America, Desmodus rotundus, the common vampire bat, one of three sanguivorous species in the family Phyllostomidae, is one of the wildlife species responsible for the transmission of the rabies virus to other domestic animals and humans (Meske et al. 2021). Being a homeothermic species, D. rotundus exhibits a varied response to extreme temperatures in laboratory settings (Lyman and Wimsatt 1966). Canine rabies has been effectively controlled in Brazil, yet bovine rabies is still endemic because of the presence of vampire bats. This zoonotic viral disease causes an annual loss of 17 million US dollars (Ferreira et al. 2012). Climate conditions can affect bat population dynamics and rabies cases, especially in domestic animals and humans. Several studies have shown that factors such as temperature, climate, rainfall, and the ENSO affect the prevalence of rabies in specific areas, seasonally and throughout the years (Santos et al. 2019). Under climate change conditions, bat species may expand their range in response to rising temperatures (Ancillotto et al. 2016), and the mechanisms by which bat range shifts in response to climate change may include dependence on water, roost quality, bat physiology, and phenology (Cappelli et al. 2021). Modeling future conditions for the sanguivorous bats along the US border with Mexico, the authors showed areas in South Texas could become suitable for this species’ habitation by 2070 (Hayes and Piaggio 2018).
Climate change is a great concern for bat survival. In addition to spreading diseases among humans, climate change also affects bats themselves. Grey long-eared bats are among the UK’s rarest mammals, numbering fewer than 1000, and have migrated to the UK from Spain and Portugal as they struggle in the increasingly hot climates of these two countries. The effects of heat waves have been quite dramatic in Australia, resulting in the death of flying foxes yearly (Groc 2021). As climate change progresses and these events become more common, bat populations may face further reduction. Numerous bats dying in heat waves can lead to ecological imbalances and the spread of zoonotic diseases (Nitnaware 2023). Large numbers of sick or dead bats lying on the ground can increase the risk of infection due to increased human-animal-bat interactions, especially if members of the public try to rescue ill bats or remove the remains of dead bats. Bat mass mortality events threaten public health because of the increased risk of disease transmission from bats to humans (Paterson et al. 2014).
Damage to the natural habitat
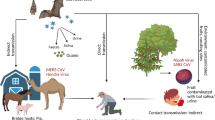
Destruction of natural habitats due to natural disasters and changes in human social structures can lead to higher disease outbreaks in wildlife, posing a significant risk to humans through zoonotic spillover (Faust et al. 2018). In tropical Asia, 65% of the main forest habitat has been lost, with specifically high rates of destruction recorded for Bangladesh (96%), Sri Lanka (86%), India (78%), and Vietnam (76%) (Primack and Morrison 2013). Loss of natural habitats of bats may lead to outbreaks of bat-associated viral infections in humans and animals (Chattu et al. 2018). As humans encroach on the environment, many bats are dislocated from natural foraging grounds, which can reduce their numbers. Bats, in turn, adjust themselves in human-made rural or urban constructions. However, as the bats adapt to human habitation, it becomes more likely that bats will have increased chances of human contact. Pteropus sp., the fruit bat, has been identified as a natural host of the Nipah virus. Sixty-five Pteropus species are distributed from Madagascar island of Africa through the Indian subcontinent to Southeast Asia and Australia and as far east as the Cook Islands. Some Pteropus species are the largest of all bats, exhibiting a weight of 1.2 kg and a wingspan of up to 1.7 m. (Neuweiler 2000). Since bats are known to be migratory or travel long distances (e.g.>200 km during migration, Krauel and McCracken 2013), they have the potential to transfer zoonotic viruses between countries through their movements (Breed et al. 2010). In Australia, all bat-related viruses, such as Hendra, Menangle, and Australian bat lyssavirus that have emerged are hypothesized to be linked to habitat loss caused by deforestation and agricultural reinforcement (Jones et al. 2013). Nipah virus (NiV) outbreaks will likely continue in affected countries due to the large distribution of native fruit bats in South and Southeast Asia. Flying foxes become stressed and starved as their habitat is uprooted by human activity and their immune system weakens. Because of the weakened immune system, their viral load increases, and they shed more viruses in their urine and saliva (Halpin et al. 2000). Similar fluctuations in virus shedding may be associated with physiological states of stress or seasonality. Among all other South Asian countries, Bangladesh is considered an important global hotspot for zoonotic spillover to humans (Allen et al. 2017). The government of Bangladesh has selected six diseases through a one-health zoonotic disease priority workshop, including three bat-borne diseases, which were Nipah virus, rabies, and zoonotic influenza (CDC 2017). Bangladesh alone contributes about 42% of the global Nipah disease burden and recorded 325 human cases from 2001 to 2022, with a case fatality rate of 71% (Pillai et al. 2020). A total of six outbreaks of NiV have been reported in India. The disease first broke out in the West Bengal state bordering Bangladesh in 2001 and 2007, during which seventy-one cases were reported, with 50 deaths between the outbreaks. Four other outbreaks have occurred in the country’s southern district of Kerala in recent years. The virus found in Kerala has been identified as the Indian genotype or I-genotype by the National Institute of Virology, Pune, identified to be identical to the NiV strain found in Bangladesh (World Health Organization 2023b). According to the World Health Organization (2023b), the Kerala outbreak strain originated in Bangladesh in 2001. It often causes small outbreaks with high mortality. To reach Kerala, the virus must have spread undetected from Bangladesh or the neighboring Indian state of West Bengal over more than 2,000 km. Many Indian states and some countries like Sri Lanka, southern China, Nepal, Bhutan, Myanmar, Thailand, and Laos are within the same range of distance from Bangladesh and have significant populations of fruit bats (Jayanth 2023). In Malaysia and Singapore, the infection was initially detected among workers involved in pig slaughter in 1998–1999 (Fig. 5). The virus spread in Malaysia due to the mismanagement of large piggeries and unplanned pulpwood deforestation, the natural habitat of NiV-carrying bats (Halder and Chakravarty 2006). Also, Malaysia lost 14.4% of its forest cover from 2000 to 2012, at the rate of one football pitch every 1.5 min; it lost 4.5 million hectares of its dense forest (Rana and Singh 2015) (Fig. 5). Similarly, the mode of transmission was observed during the outbreak on the island of Mindanao, Philippines, in persons involved in the slaughter of infected horses and consumption of infected meat (Paton et al. 1999). First in Bangladesh and then in India, NiV spillover occurred during the consumption of raw or fermented palm fruit juice contaminated with saliva, urine, and feces of bats (Hughes et al. 2009). Date palm juice consumption is popular in several Southeast Asian countries, including India, Bangladesh, Indonesia, Malaysia, Thailand, and the Philippines. Due to urbanization and increasing habitat destruction, bats progressively find homes in human habitats, leading to human-bat disputes (Hassan et al. 2020). Pteropus bats roost on trees in hundreds at the time of day and forage for fruits, pollen, and nectar at night. The roosting trees are usually located on the side of water bodies (Dey et al. 2013). Mature mangrove forests could offer distinctive roosting opportunities for bats because they support a high density of habitat trees, a stable microclimate, and a possibly low abundance of competitors and predators (McConville et al. 2013). About 46% of the world’s mangroves are present in South Asia, Southeast Asia, and the Asia-Pacific sub-region (Giri et al. 2011). However, the deforestation rate in Indonesia due to land conversion for shrimp farming and settlement ranges from 182,091 to 2,000,000 hectares annually (Arifanti et al. 2021). Similar trends are noticed in Southeast Asian countries such as Myanmar, Vietnam, and Thailand (Luo and Chui 2022). The Philippines lost 10.5% of mangroves between 1990 and 2010 (Long et al. 2014), while Cambodia experienced a 42% reduction from 1989 to 2017 (Veettil and Quang 2019). Although there has been no evidence of NiV infection in Cambodia, Cappelle et al. (2020) confirmed that NiV is prevalent in Cambodian fruit bats. Deforestation is not only caused by shrimp farming but also by the extension of palm tree plantations on forest lands, especially in Indonesia and Malaysia (Richards and Friess 2016). The Sundarbans, the world’s largest mangrove ecosystem in the sub-region, covers about 1,000,000 hectares of area at the India-Bangladesh interface (Quader et al. 2017). The most significant loss of mangroves has occurred in Bangladesh, where the initial loss rate was 80% from 2010 to 2016 (Goldberg et al. 2020). Across the sub-region, mangrove loss is caused by land-cover conversion, over-harvesting, and pollution (Giri et al. 2008). Chattu et al. (2018) mention that in Bangladesh urbanization and deforestation have contributed to immense overlap between human and bat habitats in some areas. Even in India, a large part of the mangrove forest has been destroyed due to increased aquaculture. In India, about 40% of the mangrove habitat along the western coastline has been converted for aquaculture (Kathiresan 2022). Bat populations in many Indian states have serological evidence of exposure to NiV. Many places in South and Southeast Asia have NiV reservoirs which may cause disease outbreaks in different regions in the future (Jayanth 2023). An overview of the yearly distribution of Nipah cases in humans worldwide is shown in Table 2, along with the case fatality rates.
Schematic representation of the Nipah virus outbreak in Malaysia. As a result of habitat destruction and human area expansion, fruit bats have migrated to localities in search of habitat and food in Malaysia. Because pig farms were located near fruit trees, domestic pigs became infected through exposure to bat contaminants, particularly by eating bat-eaten fruit or through contact with bat urine. NiV was then transmitted from pigs to humans through direct contact
The common vampire bat (D. rotundus), a hematophagous bat, is one of the main sylvatic rabies reservoirs and source of human and livestock infection in Latin America (Bergner et al. 2020). Common vampire bats range throughout South America (except south of the Patagonia region) to northern Mexico (Bergner et al. 2020). Human activities have contributed to increasing the density and distribution of the common vampire bat through an abundant food supply (cattle ranching) and deforestation of large areas. The deforestation of large areas, which represents an increasing problem in many South American countries, promotes the dispersal of vampire bats in search of a new location to colonize and may favor the contact between humans, livestock, and bats (Gupta 2005; Botto Nuñez et al. 2020). It has been proposed that bats’ tolerance of fragmentation and habitat loss may be connected to their ability to cross open areas to reach other vegetation or forest fragments and use resources within the matrix (Schulze et al. 2000). Brazil is the fifth largest country by territorial expansion and has the largest cattle stock in Latin America (OECD/FAO 2019), with livestock production important for national and international trade. Deforestation has occurred in some biomes in Brazil, particularly in the Amazon, where extensive cattle ranching has increased pressure on large forests, significantly altering the landscape (Dos Santos et al. 2022). The abysmal lack of strategic planning for expanding areas earmarked for agriculture and livestock alters the landscape and exacerbates the risk of outbreaks of infectious diseases among animals and humans (Fig. 6). Though in recent times incidences of rabies in humans and animals have reduced, still rabies is a potential disease for the emergence of outbreaks because vampire bats have a great capacity to adapt to anthropogenic environmental changes (Gomes et al. 2010). Coupled with this, the ability of vampire bats to travel great distances, greater than 10 km, and use all types of forest to traverse large agricultural expansions (Medina et al. 2007) makes rabies a disease capable of crossing states and countries. Botto Nuñez et al. (2020) noted that the Uruguay-Brazil border is most likely the source of the rabies epidemics in cattle that began in 2007. During the last decade in Brazil, rabies has been transmitted to humans through bites by D. rotundus bats (52.60%), dogs (23.70%), non-human primates (10.50%), and felines (10.50%) (Ministry of Health 2024) and there have been an increasing number of reports of D. rotundus bites on humans, posing a potential threat to public health in this country (Benavides et al. 2020). D. rotundus is the main transmitter of rabies in livestock animals in Brazil (100.00%, n = 50 and 99.20%, n = 650) (Kobayashi et al. 2008; Pimentel et al. 2022). Cases of rabies in livestock animals are frequently reported, with cattle being the most affected among the livestock animals (Mello et al. 2019). Vampire bats probably prefer cattle because cattle populations are constantly present and exist in higher densities than native prey mammals (Becker et al. 2018). Furthermore, the incidence of this disease has recently been on the rise in equines in Brazil (Oliveira et al. 2022).
The figure demonstrates how to prevent the interspecies transmission of pathogens, thereby reducing the risk of human outbreaks caused by bats. As the next pandemic is likely caused by a pathogen without human exposure, the highest priority should be to preserve the ecosystem and improve its resilience through restoration. In areas where humans and reservoir hosts share the landscape, priority should be given to protecting the area necessary for reservoir hosts to feed, socialize, and rest. At the same time, protecting human communities and livestock from exposure to zoonotic pathogens is needed
Consumption of bushmeat and the role of wet markets
Tanalgo et al. (2023) found that 19% (N = 254 spp.) of 1320 bats on the IUCN red list were hunted. Bat hunting is widespread, especially for large and colony-bound species, which are easy targets for hunting. In addition to habitat loss, hunting is a major threat to many bat species (Fritz et al. 2009). Bat hunting often involves various socioeconomic and cultural motivations, but food security is a major factor in rural areas (Oedin et al. 2021). In tropical regions of Africa and Asia, bats are hunted and consumed, especially fruit bats (Ghosh 2020). Between 2014 and 2016, about 28,616 people were infected, and 11,310 died in several West African countries during the Ebola virus outbreak, where bushmeat consumption was a transmission route (Wirsiy et al. 2021). Baudel et al. (2019) conducted a study of 135 people in southern Cameroon, where direct human contact with bats was found to be substantial, with 40% of respondents reporting consuming bats, 28% hunting them, 22% informed that children catch them, and 17% responded being previously bitten by bats. Hunting and consumption of bushmeat by humans carry considerable risk for cross-species disease transmission (Santos et al. 2019), particularly where bats are included. More recently, an outbreak of Sudan ebolavirus in Uganda infected 160 people between September 2022 and January 2023 and killed 77 people before it was brought under control (Okamoto et al. 2022). Uganda has several areas with high bat densities, some of which have experienced outbreaks of hemorrhagic fevers due to unknown pathogens that have been most probably transmitted by bats (MacNeil et al. 2010). A study identified that hunting and consuming bats was the top activity during an outbreak (Ninsiima et al. 2024). Respondents who hunted were ten times more likely (as a risk factor) than farmers to be exposed to bats (Vora et al. 2020). Bats are hunted for food in many parts of the world and can pose a risk to human health by spreading zoonotic pathogens from bats to humans (Vora et al. 2020). Bat hunters have domestic animals at home, where they slaughter and prepare bushmeat in front of them. Sometimes, they keep the bats alive and throw away the bat remains or feed them to pets, increasing the risk of disease transmission from bats to animals and humans. A human NiV outbreak occurred in Malaysia when bats infected pigs and the pigs infected humans (Nahar et al. 2020). Hunting, consuming, and trading bats put humans in direct contact with zoonotic pathogens that the animals may harbor, thus increasing the potential for spillover. Those involved in hunting, butchering, and consuming wildlife are at risk of infection by close contact, such as through mucosal and transcutaneous routes with live and dead animals or via contaminated sources, for example, fomites and feces (Cantlay et al. 2017). Lyssaviruses should be considered a greater infection risk for hunters because cases of fatal encephalitis from bat bites and scratches have occurred in Australia (ProMED-mail 2014). Openshaw et al. (2017) found that bat hunting occurred in 49% of the villages surveyed in Bangladesh and that bat hunting may increase the risk of NiV infection.
Live and wet markets are dense concentrations of large numbers of species that may never encounter each other in natural conditions and that may have never interacted in their evolutionary history because they do not share the same ecosystem (Lynteris 2016; Naguib et al. 2021; Galindo-González 2022). Various wild species are available in wholesale markets in Africa and Asia, either dead, frozen, or alive. In most cases, bats, rats, snakes, insects, sea urchins, turtles, sea cucumbers, civets, hedgehogs, and groundhogs are poached and sold illegally (Lynteris 2016; Zhang and Holmes 2020; Zheng 2020; Naguib et al. 2021). The study showed that bat-related diseases could spread through livestock markets (Naguib et al. 2021; Galindo-González 2022). For example, the Ebola virus in West and Central Africa, the Marburg virus in sub-Saharan Africa, and the Nipah virus in South and Southeast Asia can spread through live exotic animals or bush meat consumption (Marí Saéz et al. 2015; Judson and Munster 2023). Emerging infectious disease risks arise from interactions between humans, food products, live domestic animals for sale, and wild and scavenging animals (Naguib et al. 2021; Galindo-González 2024). Moreover, these animals interact with domestic species like pigs, rabbits, ducks, cats, chickens, dogs, etc., and humans, leading to cross-species transmission (Tian et al. 2022; Salinas-Ramos et al. 2021). Southeast Asia and China are global hotspots of emerging zoonotic diseases (Allen et al. 2017). In these regions, wildlife is generally sold in open markets, and such markets have been in the spotlight as a source of zoonotic viruses. For example, it is believed that China’s wet market in Wuhan played a crucial role in the initial spread of COVID-19 (Xiao et al. 2021). At that time, China immediately shut these markets over fears of coronavirus propagation (Xiao et al. 2021). Even Siamese Crocodiles, Indian Peafowls, Common Pheasants, raccoon dogs, Foxes, and Amur hedgehogs are used in the market as food and kept in cramped, dirty conditions like market cages. According to the World Economic Forum (Beech 2020), “it was not a wet market for China strictly, but a wildlife market”. The disease can spread easily through body fluids to handlers and customers. Indonesia’s wildlife markets are like a cafeteria for animal pathogens (Paddock and Sijabat 2020), Morcatty et al. (2022) reported that short-nosed fruit bats and large flying foxes are also sold in large quantities in ten out of fourteen markets.
Future perspective
The chances of the emergence of infectious disease increase in high-biodiversity areas experiencing changes in land use (Allen et al. 2017) and in degraded landscapes (Bloomfield et al. 2020). Well-implemented area-based management can reduce land-use change (Bruner et al. 2001). Unprotected areas with high biodiversity or important habitat features at risk of land use change must be identified and protected, particularly where bat habitats exist. To prevent range expansion into urban and semi-urban areas, natural bat habitats must be better protected to allow adequate space for bat populations, where the risk of spillover may increase through contact with humans and livestock (Schneeberger and Voigt 2016) (Fig. 6).
To avoid future epidemics of zoonotic diseases, governments must rigorously execute bans on the sale and trade of wildlife for food, and they must adhere to food safety and hygienic conditions ( https://www.bbc.com/news/world-australia-52391783 ). National policies to protect or control bat hunting are either absent or lack enforcement in many countries (Oedin et al. 2021 ). More measures may be needed to address illegal hunting (Geldmann et al. 2019 ). Regardless of the public’s negative perception of bats, they are important components of all terrestrial biological communities (Del Vaglio et al. 2011 ). They help control insects, harvest forest seeds or aid in seed dispersal, enhance soil fertility and nutrient distribution, recycle and pollinate plants that provide food for humans and other species, and act as predators of insects (Russo et al. 2018; Enríquez-Acevedo et al. 2020 ). In pest control, they provide at least $3.7 billion in services each year in North America alone (Boyles et al. 2011 ). Ramírez-Fráncel et al. ( 2022 ) identified 409 bat species that provide ecosystem services, 549 plant species are pollinated or dispersed by bats, and bats eat 752 insect species. Resolving the bat-human conflict requires rigorous approaches to underpin human behavior and develop effective interventions to promote behavioral change (Kingston 2016 ). Integrating local hunter/collector communities (e.g. vendors and customers) and Indigenous peoples into early warning systems for disease outbreaks that link to national and international public health reporting systems. Community engagement activities should co-design interventions and conduct them to strengthen the perceived fairness and legitimacy of new laws or standards. Public awareness should be created about the risks of contact with wildlife and the increased chances of transmission of diseases associated with wildlife hunting. The importance of the beneficial ecosystem services bats provide should also be highlighted in public awareness programs. Social acknowledgment campaigns can help influence public behavior for the benefit of biodiversity and society (Salazar et al. 2019 ). It should be noted that appropriate conservation measures can even reduce the risk of viral spread from bat populations to human populations (Schneeberger and Voigt 2016; Weber et al. 2023 ).
Conclusion
Through millions of years of evolution, the bat population on earth has adapted their immune system so that they do not suffer from the deadly outcomes of different microbes, especially viruses. However, they can still act as a carrier for several viral diseases. Bats have adjusted to various niches and habitats, and confrontation with humans is generally avoided. This has also limited the potential of outbreaks of deadly diseases from bats to humans. However, with global warming, changing climate, bat habitat destruction, and bushmeat consumption, the frequency of bat-human contact has increased by multiple folds. The anthropogenic factors have also influenced the change and shift of the bat population. This has further increased the chances of frequent bat-human contact. This increase in contact between bats and humans has also increased the chances of an outbreak and the consecutive spread of zoonotic diseases, which were suppressed in nature. Governments of the world and people, in general, should consider the necessary steps that can be taken to conserve the bat population in their natural habitats and to minimize frequent contact of humans with this vital fauna. If serious efforts are not implemented soon to prevent global warming, climate change, and bush meat consumption, it might be too late to control newer microbial outbreaks.
Data availability
No datasets were generated or analysed during the current study.
References
Ahn M, Cui J, Irving AT, Wang LF (2016) Unique loss of the PYHIN gene family in bats amongst mammals: implications for inflammasome sensing. Sci Rep 6(1):21722. https://doi.org/10.1038/srep21722
Ahn M, Anderson DE, Zhang Q, Tan CW, Lim BL, Luko K, Wen M, Chia WN, Mani S, Wang LC, Ng JH (2019) Dampened NLRP3-mediated inflammation in bats and implications for a special viral reservoir host. Nat Microbiol 4(5):789–799. https://doi.org/10.1038/s41564-019-0371-3
Allen T, Murray KA, Zambrana-Torrelio C, Morse SS, Rondinini C, Di Marco M, Breit N, Olival KJ, Daszak P (2017) Global hotspots and correlates of emerging zoonotic diseases. Nat Commun 8(1):1124. https://doi.org/10.1038/s41467-017-00923-8
Amman BR, Nyakarahuka L, McElroy AK, Dodd KA, Sealy TK, Schuh AJ, Shoemaker TR, Balinandi S, Atimnedi P, Kaboyo W, Nichol ST (2014) Marburgvirus resurgence in Kitaka mine Bat population after extermination attempts. Uganda Emerg Infect Dis 20(10):1761. https://doi.org/10.3201/eid2010.140696
Ancillotto L, Santini L, Ranc N, Maiorano L, Russo D (2016) Extraordinary range expansion in a common bat: the potential roles of climate change and urbanisation. Sci Nat 103:1–8. https://doi.org/10.1007/s00114-016-1334-7
Anyamba A, Chretien JP, Britch SC, Soebiyanto RP, Small JL, Jepsen R, Forshey BM, Sanchez JL, Smith RD, Harris R, Tucker CJ (2019) Global disease outbreaks associated with the 2015–2016 El Niño event. Sci Rep 9(1):1930. https://doi.org/10.1038/s41598-018-38034-z
Arifanti VB, Novita N, Tosiani A (2021) Mangrove deforestation and CO2 emissions in Indonesia. InIOP Conference Series: Earth and Environmental Science 874(1):012006. https://doi.org/10.1088/1755-1315/874/1/012006
Baid K, Irving AT, Jouvenet N, Banerjee A (2024) The translational potential of studying Bat immunity. Trends Immunol 45(3):188–197. https://doi.org/10.1016/j.it.2024.01.007
Baker ML, Zhou P, Bat (2015) immunology Bats and viruses: a new frontier of emerging infectious diseases 2:327– 48. https://doi.org/10.1002/9781118818824.ch14
Banerjee A, Rapin N, Bollinger T, Misra V (2017) Lack of inflammatory gene expression in bats: a unique role for a transcription repressor. Sci Rep 7(1):2232. https://doi.org/10.1038/s41598-017-01513-w
Banerjee A, Zhang X, Yip A, Schulz KS, Irving AT, Bowdish D, Golding B, Wang LF, Mossman K (2020) Positive selection of a Serine residue in Bat IRF3 confers enhanced antiviral protection. Iscience 23(3). https://doi.org/10.1016/j.isci.2020.100958
Barton ES, White DW, Cathelyn JS, Brett-McClellan KA, Engle M, Diamond MS, Miller VL, Virgin IVHW (2007) Herpesvirus latency confers symbiotic protection from bacterial infection. Nature 447(7142):326–329. https://doi.org/10.1038/nature05762
Baudel H, De Nys H, Mpoudi Ngole E, Peeters M, Desclaux A (2019) Understanding Ebola virus and other zoonotic transmission risks through human–bat contacts: exploratory study on knowledge, attitudes and practices in Southern Cameroon. Zoonoses Public Health 66(3):288–295. https://doi.org/10.1111/zph.12563
Becker DJ, Czirják GÁ, Volokhov DV, Bentz AB, Carrera JE, Camus MS, Navara KJ, Chizhikov VE, Fenton MB, Simmons NB, Recuenco SE (2018) Livestock abundance predicts vampire Bat demography, immune profiles and bacterial infection risk. Philosophical Trans Royal Soc B: Biol Sci 373(1745):20170089. https://doi.org/10.1098/rstb.2017.0089
Beech P (2020) What we’ve got wrong about china’s ‘wet markets’ and their link to COVID-19. InWorld Economic Forum:18.https://www.weforum.org/agenda/2020/04/china-wet-marketscovid19-coronavirus-explained/
Beere HM, Wolf BB, Cain K, Mosser DD, Mahboubi A, Kuwana T, Tailor P, Morimoto RI, Cohen GM, Green DR (2000) Heat-shock protein 70 inhibits apoptosis by preventing recruitment of procaspase-9 to the Apaf-1 apoptosome. Nat Cell Biol 2(8):469–475. https://doi.org/10.1038/35019501
Benavides JA, Megid J, Campos A, Hampson K (2020) Using surveillance of animal bite patients to Decipher potential risks of rabies exposure from domestic animals and wildlife in Brazil. Front Public Health 8:318. https://doi.org/10.3389/fpubh.2020.00318
Bergner LM, Orton RJ, Benavides JA, Becker DJ, Tello C, Biek R, Streicker DG (2020) Demographic and environmental drivers of metagenomic viral diversity in vampire bats. Mol Ecol 29(1):26–39. https://doi.org/10.1111/mec.15250
Bloomfield LS, McIntosh TL, Lambin EF (2020) Habitat fragmentation, livelihood behaviors, and contact between people and nonhuman primates in Africa. Landscape Ecol 35(4):985–1000. https://doi.org/10.1007/s10980-020-00995-w
Boardman WS, Baker ML, Boyd V, Crameri G, Peck GR, Reardon T, Smith IG, Caraguel CG, Prowse TA (2020) Seroprevalence of three paramyxoviruses; Hendra virus, Tioman virus, Cedar virus and a rhabdovirus, Australian Bat lyssavirus, in a range expanding fruit Bat, the Grey-headed flying Fox (Pteropus poliocephalus). PLoS ONE 15(5):e0232339. https://doi.org/10.1371/journal.pone.0232339
Botto Nunez G, Becker DJ, Lawrence RL, Plowright RK (2020) Synergistic effects of grassland fragmentation and temperature on bovine rabies emergence. EcoHealth 17:203–216. https://doi.org/10.1007/s10393-020-01486-9
Boyles JG, Cryan PM, McCracken GF, Kunz TH (2011) Economic importance of bats in agriculture. Science 332(6025):41–42. https://doi.org/10.1126/science.1201366
Breed AC, Field HE, Smith CS, Edmonston J, Meers J (2010) Bats without borders: long-distance movements and implications for disease risk management. EcoHealth 7:204–212. https://doi.org/10.1007/s10393-010-0332-z
British Broadcasting Corporation (2020) Coronavirus: Australia urges G20 action on wildlife wet markets. British Broadcasting Corporation website. https://www.bbc.com/news/world-australia-52391783
Brook CE, Dobson AP (2015) Bats as ‘special’reservoirs for emerging zoonotic pathogens. Trends Microbiol 23(3):172–180. https://doi.org/10.1016/j.tim.2014.12.004
Bruner AG, Gullison RE, Rice RE, Da Fonseca GA (2001) Effectiveness of parks in protecting tropical biodiversity. Science 291(5501):125–128. https://doi.org/10.1126/science.291.5501.125
Burgin CJ, Colella JP, Kahn PL, Upham NS (2018) How many species of mammals are there? J Mammal 99(1):1–4. https://doi.org/10.1093/jmammal/gyx147
Cadet J, Wagner JR (2013) DNA base damage by reactive oxygen species, oxidizing agents, and UV radiation. Cold Spring Harb Perspect Biol 5(2):a012559. https://doi.org/10.1101/cshperspect.a012559
Calisher CH, Childs JE, Field HE, Holmes KV, Schountz T (2006) Bats: important reservoir hosts of emerging viruses. Clin Microbiol Rev 19(3):531–545. https://doi.org/10.1128/cmr.00017-06
Cantlay JC, Ingram DJ, Meredith AL (2017) A review of zoonotic infection risks associated with the wild meat trade in Malaysia. EcoHealth 14:361–388. https://doi.org/10.1007/s10393-017-1229-x
Cappelle J, Hoem T, Hul V, Furey N, Nguon K, Prigent S, Dupon L, Ken S, Neung C, Hok V, Pring L (2020) Nipah virus circulation at human–bat interfaces. Cambodia Bull World Health Organ 98(8):539. https://doi.org/10.2471/BLT.20.254227
Cappelli F, Costantini V, Consoli D (2021) The trap of climate change-induced natural disasters and inequality. Glob Environ Change 70:102329. https://doi.org/10.1016/j.gloenvcha.2021.102329
Carlson CJ, Albery GF, Merow C, Trisos CH, Zipfel CM, Eskew EA, Olival KJ, Ross N, Bansal S (2022) Climate change increases cross-species viral transmission risk. Nature 607(7919):555–562. https://doi.org/10.1038/s41586-022-04788-w
Centers for Disease Control and Prevention (CDC) (2017) One Health Zoonotic Disease Prioritization for Multisectoral Engagement in Bangladesh
Chadha MS, Comer JA, Lowe L, Rota PA, Rollin PE, Bellini WJ, Ksiazek TG, Mishra AC (2006) Nipah virus-associated encephalitis outbreak, siliguri, India. Emerg Infect Dis 12(2):235. https://doi.org/10.3201/eid1202.051247
Channappanavar R, Perlman S (2017) Pathogenic human coronavirus infections: causes and consequences of cytokine storm and immunopathology. InSeminars Immunopathol 39:529–539 Springer Berlin Heidelberg. https://doi.org/10.1007/s00281-017-0629-x
Chant K, Chan R, Smith M, Dwyer DE, Kirkland P (1998) Probable human infection with a newly described virus in the family paramyxoviridae. The NSW expert group. Emerg Infect Dis 4(2):273. https://doi.org/10.3201/eid0402.980215
Chattu VK, Kumar R, Kumary S, Kajal F, David JK (2018) Nipah virus epidemic in Southern India and emphasizing one health approach to ensure global health security. J Family Med Prim Care 7(2):275–283. https://doi.org/10.4103/jfmpc.jfmpc_137_18.who
Ching PK, de Los Reyes VC, Sucaldito MN, Tayag E, Columna-Vingno AB, Malbas FF Jr, Bolo GC Jr, Sejvar JJ, Eagles D, Playford G, Dueger E (2015) Outbreak of henipavirus infection, Philippines, 2014. Emerging infectious diseases 21(2):328. https://doi.org/10.3201/eid2102.141433
Chua KB, Koh CL, Hooi PS, Wee KF, Khong JH, Chua BH, Chan YP, Lim ME, Lam SK (2002) Isolation of Nipah virus from Malaysian Island flying-foxes. Microbes Infect 4(2):145–151. https://doi.org/10.1016/S1286-4579(01)01522-2
Chua KB, Voon K, Crameri G, Tan HS, Rosli J, McEachern JA, Suluraju S, Yu M, Wang LF (2008) Identification and characterization of a new orthoreovirus from patients with acute respiratory infections. PLoS ONE 3(11):e3803. https://doi.org/10.1371/journal.pone.0003803
Clayton E, Munir M (2020) Fundamental characteristics of Bat interferon systems. Front Cell Infect Microbiol 10:527921. https://doi.org/10.3389/fcimb.2020.527921
Cohn R (2020) Spillover warning: how we can prevent the next pandemic. Yale environment 360. How We Can Prevent the Next Pandemic - Yale E360, Spillover Warning
Del Vaglio MA, Nicolau H, Bosso L (2011) Feeding habits of the Egyptian fruit Bat Rousettus aegyptiacus on Cyprus island: a first assessment. https://doi.org/10.4404/hystrix-22.2-4587
Dempsey A, Bowie AG (2015) Innate immune recognition of DNA: A recent history. Virology 479:146–152. https://doi.org/10.1016/j.virol.2015.03.013
Dey S, Roy US, Chattopadhyay S (2013) Distribution and abundance of three populations of Indian flying Fox (Pteropus giganteus) from purulia district of West bengal, India. Taprobanica: J Asian Biodivers 5(1). https://doi.org/10.4038/tapro.v5i1.5664
Diengdoh VL, Ondei S, Hunt M, Brook BW (2022) Predicted impacts of climate change and extreme temperature events on the future distribution of fruit Bat species in Australia. Global Ecol Conserv 37:e02181. https://doi.org/10.1016/j.gecco.2022.e02181
Dos Santos AM, Da Silva CF, De Melo SN, de Almeida Junior PM, Bueno LF (2022) Influence of deforestation inside and outside Indigenous lands in the Brazilian Amazon biome. Reg Envriron Chang 22(2):77. https://doi.org/10.1007/s10113-022-01937-9
Enríquez-Acevedo T, Pérez-Torres J, Ruiz-Agudelo C, Suarez A (2020) Seed dispersal by fruit bats in Colombia generates ecosystem services. Agron Sustain Dev 40(6):45. https://doi.org/10.1007/s13593-020-00645-0
Epstein JH, Field HE (2015) Anthropogenic Epidemics: The Ecology of Bat-Borne Viruses and Our Role in their Emergence. Bats and viruses: a new frontier of emerging infectious diseases, pp.249– 79. https://doi.org/10.1002/9781118818824.ch10
Faust CL, McCallum HI, Bloomfield LS, Gottdenker NL, Gillespie TR, Torney CJ, Dobson AP, Plowright RK (2018) Pathogen spillover during land conversion. Ecol Lett 21(4):471–483. https://doi.org/10.1111/ele.12904
Ferreira RS, de Almeida RM, Nogueira DA, Oliveira NM, Fiorini JE (2012) Bovine rabies incidence in the state of Minas gerais/brazil, between 2002 and 2006. Arquivos Do Instituto Biológico 79:287–291. https://doi.org/10.1590/S1808-16572012000200019
Field H, McCall B, Barrett J (1999) Australian Bat lyssavirus infection in a captive juvenile black flying Fox. Emerg Infect Dis 5(3):438. https://doi.org/10.3201/eid0503.990316
Fritz SA, Bininda-Emonds OR, Purvis A (2009) Geographical variation in predictors of mammalian extinction risk: big is bad, but only in the tropics. Ecol Lett 12(6):538–549. https://doi.org/10.1111/j.1461-0248.2009.01307.x
Galindo-González J (2022) Live animal markets: identifying the origins of emerging infectious diseases. Curr Opin Environ Sci Health 25:100310. https://doi.org/10.1016/j.coesh.2021.100310
Galindo-González J (2024) Avoiding novel, unwanted interactions among species to decrease risk of zoonoses. Conserv Biol 38(3):e14232. https://doi.org/10.1111/cobi.14232
Ge XY, Li JL, Yang XL, Chmura AA, Zhu G, Epstein JH, Mazet JK, Hu B, Zhang W, Peng C, Zhang YJ (2013) Isolation and characterization of a Bat SARS-like coronavirus that uses the ACE2 receptor. Nature 503(7477):535–538. https://doi.org/10.1038/nature12711
Geldmann J, Manica A, Burgess ND, Coad L, Balmford A (2019) A global-level assessment of the effectiveness of protected areas at resisting anthropogenic pressures. Proceedings of the National Academy of Sciences 116(46):23209-15. https://doi.org/10.1073/pnas.1908221116
George DB, Webb CT, Farnsworth ML, O’Shea TJ, Bowen RA, Smith DL, Stanley TR, Ellison LE, Rupprecht CE (2011) Host and viral ecology determine bat rabies seasonality and maintenance. Proceedings of the National Academy of Sciences 108(25):10208-13. https://doi.org/10.1073/pnas.1010875108
Gerow CM, Rapin N, Voordouw MJ, Elliot M, Misra V, Subudhi S (2019) Arousal from hibernation and reactivation of Eptesicus fuscus gammaherpesvirus (Ef HV) in big brown bats. Transbound Emerg Dis 66(2):1054–1062. https://doi.org/10.1111/tbed.13102
Ghosh S (2020) Bats not the enemy in the fight against COVID-19. MONGABAY. Bats not the enemy in the fight against COVID-19 (mongabay.com)
Giles JR, Eby P, Parry H, Peel AJ, Plowright RK, Westcott DA, McCallum H (2018) Environmental drivers of Spatiotemporal foraging intensity in fruit bats and implications for Hendra virus ecology. Sci Rep 8(1):9555. https://doi.org/10.1038/s41598-018-27859-3
Giri C, Zhu Z, Tieszen LL, Singh A, Gillette S, Kelmelis JA (2008) Mangrove forest distributions and dynamics (1975–2005) of the tsunami-affected region of Asia. J Biogeogr 35(3):519–528. https://doi.org/10.1111/j.1365-2699.2007.01806.x
Giri C, Ochieng E, Tieszen LL, Zhu Z, Singh A, Loveland T, Masek J, Duke N (2011) Glob Ecol Biogeogr 20(1):154–159. https://doi.org/10.1111/j.1466-8238.2010.00584.x. Status and distribution of mangrove forests of the world using earth observation satellite data
Goldberg L, Lagomasino D, Thomas N, Fatoyinbo T (2020) Global declines in human-driven Mangrove loss. Glob Change Biol 26(10):5844–5855. https://doi.org/10.1111/gcb.15275
Gomes MN, Monteiro AM, Lewis N, Gonçalves CA, de Souza Nogueira Filho V (2010) Landscape risk factors for attacks of vampire bats on cattle in Sao paulo, Brazil. Prev Vet Med 93(2–3):139–146. https://doi.org/10.1016/j.prevetmed.2009.10.006
Gong L, Li J, Zhou Q, Xu Z, Chen L, Zhang Y, Xue C, Wen Z, Cao Y (2017) A new bat-HKU2–like coronavirus in swine, china, 2017. Emerg Infect Dis 23(9):1607. https://doi.org/10.3201/eid2309.170915
Groc I (2021) Hot houses: the race to save bats from overheating as temperatures rise. The Guardian. Hot houses: the race to save bats from overheating as temperatures rise| Wildlife| The Guardian
Gupta R (2005) Recent outbreak of rabies infections in Brazil transmitted by vampire bats. Weekly releases (1997–2007). 10(45):2831. https://doi.org/10.2807/esw.10.45.02831-en
Guth S, Hanley KA, Althouse BM, Boots M (2020) Ecological processes underlying the emergence of novel enzootic cycles: arboviruses in the neotropics as a case study. PLoS Negl Trop Dis 14(8):e0008338. https://doi.org/10.1371/journal.pntd.0008338
Haddaway NR, Page MJ, Pritchard CC, McGuinness LA (2022) PRISMA2020: an R package and Shiny app for producing PRISMA 2020-compliant flow diagrams, with interactivity for optimised digital transparency and open synthesis. Campbell Syst Reviews 18(2):e1230. https://doi.org/10.1002/cl2.1230
Halder A, Chakravarty A (2006) Nipah virus encephalitis: A cause for concern for Indian neurologists? Ann Indian Acad Neurol 9(3):137–144. https://doi.org/10.4103/0972-2327.27655
Halpin K, Young PL, Field HE, Mackenzie JS (2000) Isolation of Hendra virus from pteropid bats: a natural reservoir of Hendra virus. J Gen Virol 81(8):1927–1932. https://doi.org/10.1099/0022-1317-81-8-1927
Han HJ, Wen HL, Zhou CM, Chen FF, Luo LM, Liu JW, Yu XJ (2015) Bats as reservoirs of severe emerging infectious diseases. Virus Res 205:1–6. https://doi.org/10.1016/j.virusres.2015.05.006
Hanley DE, Bourassa MA, O’Brien JJ, Smith SR, Spade ER (2003) A quantitative evaluation of ENSO indices. J Clim 16(8):1249–1258. https://doi.org/10.1175/1520-0442(2003)16%3C;1249:AQEOEI%3E;2.0.CO;2
Hassan MM, Kalam MA, Alam M, Shano S, Faruq AA, Hossain MS, Islam MN, Khan SA, Islam A (2020) Understanding the community perceptions and knowledge of bats and transmission of Nipah virus in Bangladesh. Animals 10(10):1814. https://doi.org/10.3390/ani10101814
Hayes MA, Piaggio AJ (2018) Assessing the potential impacts of a changing climate on the distribution of a rabies virus vector. PLoS ONE 13(2):e0192887. https://doi.org/10.1371/journal.pone.0192887
Hölzer M, Schoen A, Wulle J, Müller MA, Drosten C, Marz M, Weber F (2019) Virus-and interferon alpha-induced transcriptomes of cells from the microbat Myotis daubentonii. Iscience 19:647–661. https://doi.org/10.1016/j.isci.2019.08.016
Huang C, Wang Y, Li X, Ren L, Zhao J, Hu Y, Zhang L, Fan G, Xu J, Gu X, Cheng Z (2020) Clinical features of patients infected with 2019 novel coronavirus in wuhan, China. Lancet 395(10223):497–506. https://doi.org/10.1016/S0140-6736(20)30183-5
Hughes JM, Wilson ME, Luby SP, Gurley ES, Hossain MJ (2009) Transmission of human infection with Nipah virus. Clin Infect Dis 49(11):1743–1748. https://doi.org/10.1086/647951
IEDCR, National Bulletin of Public Health (2022) An Update on Nipah Encephalitis Surveillance 3(3). 2023-01-27-18-04-1e9ef6ab012f87112ad43e5b322e9820.pdf (iedcr.gov.bd).
Irving AT, Ahn M, Goh G, Anderson DE, Wang LF (2021) Lessons from the host defences of bats, a unique viral reservoir. Nature 589(7842):363–370. https://doi.org/10.1038/s41586-020-03128-0
Jayanth AS (2023) Nipah infection possible in more places in Southeast Asia, says Nature article. The Hindu. Nipah infection possible in more places in Southeast Asia, says Nature article - The Hindu
Jebb D, Foley NM, Whelan CV, Touzalin F, Puechmaille SJ, Teeling EC (2018) Population level mitogenomics of long-lived bats reveals dynamic heteroplasmy and challenges the free radical theory of ageing. Sci Rep 8(1):13634. https://doi.org/10.1038/s41598-018-31093-2
Jennings J, Sang Y (2019) Porcine interferon complex and co-evolution with increasing viral pressure after domestication. Viruses 11(6):555. https://doi.org/10.3390/v11060555
Jones BA, Grace D, Kock R, Alonso S, Rushton J, Said MY, McKeever D, Mutua F, Young J, McDermott J, Pfeiffer DU (2013) Zoonosis emergence linked to agricultural intensification and environmental change. Proceedings of the national academy of sciences 110(21):8399– 404. https://doi.org/10.1073/pnas.1208059110
Judson SD, Munster VJ (2023) The multiple origins of Ebola disease outbreaks. J Infect Dis 228(Supplement7):S465–S473. https://doi.org/10.1093/infdis/jiad352
Kapp K, Maul J, Hostmann A, Mundt P, Preiss JC, Wenzel A, Thiel A, Zeitz M, Ullrich R, Duchmann R (2010) Modulation of systemic antigen-specific immune responses by oral antigen in humans. Eur J Immunol 40(11):3128–3137. https://doi.org/10.1002/eji.201040701
Kathiresan K (2022) Mangrove forests of india: an overview. Mangroves: Biodivers Livelihoods Conserv 233–270. https://doi.org/10.1007/978-981-19-0519-3_11
Kingston T (2016) Cute, creepy, or crispy—how values, attitudes, and norms shape human behavior toward bats. Bats Anthropocene: Conserv Bats Chang World pp571–95. 978-3-319-25220-9_18.pdf (springer.com)
Kobayashi Y, Sato G, Mochizuki N, Hirano S, Itou T, Carvalho AA, Albas A, Santos HP, Ito FH, Sakai T (2008) Molecular and geographic analyses of vampire bat-transmitted cattle rabies in central Brazil. BMC Vet Res 4:1–9. https://doi.org/10.1186/1746-6148-4-44
Koh J, Itahana Y, Mendenhall IH, Low D, Soh EX, Guo AK, Chionh YT, Wang LF, Itahana K (2019) ABCB1 protects Bat cells from DNA damage induced by genotoxic compounds. Nat Commun 10(1):2820. https://doi.org/10.1038/s41467-019-10495-4
Krauel JJ, McCracken GF (2013) Recent advances in Bat migration research. Bat Evol Ecol Conserv 293–313. https://doi.org/10.1007/978-1-4614-7397-8_15
La Cruz-Rivera D, Pamela C, Kanchwala M, Liang H, Kumar A, Wang LF, Xing C, Schoggins JW (2018) The IFN response in bats displays distinctive IFN-stimulated gene expression kinetics with atypical RNASEL induction. J Immunol 200(1):209–217. https://doi.org/10.4049/jimmunol.1701214
Laing ED, Sterling SL, Weir DL, Beauregard CR, Smith IL, Larsen SE, Wang LF, Snow AL, Schaefer BC, Broder CC (2019) Enhanced autophagy contributes to reduced viral infection in black flying Fox cells. Viruses 11(3):260. https://doi.org/10.3390/v11030260
Lam TT, Jia N, Zhang YW, Shum MH, Jiang JF, Zhu HC, Tong YG, Shi YX, Ni XB, Liao YS, Li WJ (2020) Identifying SARS-CoV-2-related coronaviruses in Malayan pangolins. Nature 583(7815):282–285. https://doi.org/10.1038/s41586-020-2169-0
Latinne A, Morand S (2022) Climate anomalies and spillover of bat-borne viral diseases in the asia–pacific region and the Arabian Peninsula. Viruses 14(5):1100. https://doi.org/10.3390/v14051100
Lee YF, McCracken GF (2002) Foraging activity and food resource use of Brazilian free-tailed bats, Tadarida brasiliensis (Molossidae). Ecoscience9(3):306– 13. https://doi.org/10.1080/11956860.2002.11682717
Li W, Shi Z, Yu M, Ren W, Smith C, Epstein JH, Wang H, Crameri G, Hu Z, Zhang H, Zhang J (2005) Bats are natural reservoirs of SARS-like coronaviruses. Science 310(5748):676–679. https://doi.org/10.1126/science.1118391
Liu X, Speranza E, Muñoz-Fontela C, Haldenby S, Rickett NY, Garcia-Dorival I, Fang Y, Hall Y, Zekeng EG, Lüdtke A, Xia D (2017) Transcriptomic signatures differentiate survival from fatal outcomes in humans infected with Ebola virus. Genome Biol 18:1–7. https://doi.org/10.1186/s13059-016-1137-3
Liu Z, Liu Q, Wang H, Yao X (2024) Severe zoonotic viruses carried by different species of bats and their regional distribution. Clin Microbiol Infect 30(2):206–210. https://doi.org/10.1016/j.cmi.2023.09.025
Long J, Napton D, Giri C, Graesser J (2014) A mapping and monitoring assessment of the philippines’ Mangrove forests from 1990 to 2010. J Coastal Res 30(2):260–271. https://doi.org/10.2112/JCOASTRES-D-13-00057.1
Luo S, Chui TF (2022) Sea-level rise predicted to reduce exotic Mangrove distribution and biomass in coastal wetlands in Southern China. J Hydrol 612:128234. https://doi.org/10.1016/j.jhydrol.2022.128234
Lyman CP, Wimsatt WA (1966) Temperature regulation in the vampire bat, Desmodus rotundus. Physiological Zoology 39(2):101-9. Temperature Regulation in the Vampire Bat, Desmodus rotundus| Physiological Zoology: Vol 39, No 2 (uchicago.edu)
Lynteris C (2016) The prophetic faculty of epidemic photography: Chinese wet markets and the imagination of the next pandemic. Visual Anthropol 29(2):118–132. https://doi.org/10.1080/08949468.2016.1131484
Mackenzie JS, Childs JE, Field HE, Wang LF, Breed AC (2016) The role of bats as reservoir hosts of emerging neuroviruses. Neurotropic Viral Infections: Volume 2: Neurotropic Retroviruses, DNA Viruses, Immunity and Transmission pp.403– 54. https://doi.org/10.1007/978-3-319-33189-8_12
MacNeil A, Farnon EC, Wamala J, Okware S, Cannon DL, Reed Z, Towner JS, Tappero JW, Lutwama J, Downing R, Nichol ST (2010) Proportion of deaths and clinical features in Bundibugyo Ebola virus infection, Uganda. Emerg Infect Dis 16(12):1969. https://doi.org/10.3201/eid1612.100627
Maina JN (2000) What it takes to fly: the structural and functional respiratory refinements in birds and bats. J Exp Biol 203(20):3045–3064. https://doi.org/10.1242/jeb.203.20.3045
Mandl JN, Schneider C, Schneider DS, Baker ML (2018) Going to Bat (s) for studies of disease tolerance. Front Immunol 9:2112. https://doi.org/10.3389/fimmu.2018.02112
Marí Saéz A, Weiss S, Nowak K, Lapeyre V, Zimmermann F, Düx A, Kühl HS, Kaba M, Regnaut S, Merkel K, Sachse A (2015) Investigating the zoonotic origin of the West African Ebola epidemic. EMBO Mol Med 7(1):17–23. https://doi.org/10.15252/emmm.201404792
McConville A, Law BS, Mahony MJ (2013) Mangroves as maternity roosts for a colony of the rare east-coast free-tailed Bat (Mormopterus norfolkensis) in south-eastern Australia. Wildl Res 40(4):318–327. https://doi.org/10.1071/WR12222
Medina A, Harvey CA, Merlo DS, Vílchez S, Hernández B (2007) Bat diversity and movement in an agricultural landscape in matiguás, Nicaragua. Biotropica 39(1):120–128. https://doi.org/10.1111/j.1744-7429.2006.00240.x
Mello AK, Brumatti RC, Neves DA, Alcântara LO, Araújo FS, Gaspar AO, Lemos RA (2019) Bovine rabies: economic loss and its mitigation through antirabies vaccination. Pesquisa Veterinária Brasileira 39:179–185. https://doi.org/10.1590/1678-5150-PVB-6201
Meske M, Fanelli A, Rocha F, Awada L, Soto PC, Mapitse N, Tizzani P (2021) Evolution of rabies in South America and inter-species dynamics (2009–2018). Trop Med Infect Disease 6(2):98. https://doi.org/10.3390/tropicalmed6020098
Ministry of Health. Secretaria de Vigilância em Saúde. Sistema Nacional de Vigilância em Saúde. Raiva: O Que é, Causas, Sintomas, Tratamento, Diagnóstico e Prevenção. Ministério da Saúde; Brasília, Brazil (2020) (Accessed 5 June 2024). Available online: http://www.saude.gov.br/saude-de-a-z/raiva/#epidemiologia
Mollentze N, Streicker DG (2020) Viral zoonotic risk is homogenous among taxonomic orders of mammalian and avian reservoir hosts. Proc Natl Acad Sci 117(17):9423–9430. https://doi.org/10.1073/pnas.1919176117
Morcatty TQ, Pereyra PE, Ardiansyah A, Imron MA, Hedger K, Campera M, Nekaris KA, Nijman V (2022) Risk of viral infectious diseases from live bats, primates, rodents and carnivores for sale in Indonesian wildlife markets. Viruses 14(12):2756. https://doi.org/10.3390/v14122756
Munster VJ, Adney DR, van Doremalen N, Brown VR, Miazgowicz KL, Milne-Price S, Bushmaker T, Rosenke R, Scott D, Hawkinson A, de Wit E (2016) Replication and shedding of MERS-CoV in Jamaican fruit bats (Artibeus jamaicensis). Sci Rep 6(1):21878. https://doi.org/10.1038/srep21878
Naguib MM, Li R, Ling J, Grace D, Nguyen-Viet H, Lindahl JF (2021) Live and wet markets: food access versus the risk of disease emergence. Trends Microbiol 29(7):573–581. https://doi.org/10.1016/j.tim.2021.02.007
Nahar N, Asaduzzaman M, Mandal UK, Rimi NA, Gurley ES, Rahman M, Garcia F, Zimicki S, Sultana R, Luby SP (2020) Hunting bats for human consumption in Bangladesh. EcoHealth 17:139–151. https://doi.org/10.1007/s10393-020-01468-x
National Oceanic and Atmospheric Administration (NOAA) National Oceanic and Atmospheric Administration website, United States government (accessed 20 May 2024). Available online: https://www.noaa.gov/
Neuweiler G (2000) The biology of bats. Oxford University Press, USA. The Biology of Bats - Gerhard Neuweiler - Google Books
Ninsiima LR, Nyakarahuka L, Kisaka S, Atuheire CG, Mugisha L, Odoch T, Romano JS, Klein J, Mor SM, Kankya C (2024) Knowledge, perceptions, and exposure to Bats in communities living around Bat roosts in Bundibugyo district, uganda: implications for viral haemorrhagic fever prevention and control. BMC Infect Dis 24(1):311. https://doi.org/10.1186/s12879-024-09162-x
Nitnaware H (2023) Bats dying of heat stroke: Experts fear mass population collapse, zoonotic diseases. Down to Earth. Bats dying of heat stroke: Experts fear mass population collapse, zoonotic diseases (downtoearth.org.in)
Norberg UM, Rayner JM (1987) Ecological morphology and flight in bats (Mammalia; Chiroptera): wing adaptations, flight performance, foraging strategy and echolocation. Philosophical Trans Royal Soc Lond B Biol Sci 316(1179):335–427. https://doi.org/10.1098/rstb.1987.0030
OECD/FAO (2019) OECD-FAO agricultural outlook 2019–2028. OECD Publishing, Paris. https://doi.org/10.1787/agr_outlook-2019-en
Oedin M, Brescia F, Millon A, Murphy BP, Palmas P, Woinarski JC, Vidal E (2021) Cats Felis catus as a threat to bats worldwide: a review of the evidence. Mammal Rev 51(3):323–337. https://doi.org/10.1111/mam.12240
Okamoto F, Nishikawa Y, Twalla Y, Nakayama T, Masai S (2022) Ebola outbreak in uganda: patient values and preferences. Lancet 400(10367):1925–1926. https://doi.org/10.1016/S0140-6736(22)02321-2
Oliveira FA, Castro RJ, de Oliveira JF, Barreto FM, Farias MP, Marinho GL, Dos Santos Soares MJ, Silva-Júnior A, Schwarz DG (2022) Geographical and Temporal spread of equine rabies in Brazil. Acta Trop 227:106302. https://doi.org/10.1016/j.actatropica.2022.106302
Olivero J, Fa JE, Farfán MÁ, Márquez AL, Real R, Juste FJ, Leendertz SA, Nasi R (2020) Human activities link fruit Bat presence to Ebola virus disease outbreaks. Mammal Rev 50(1):1–0. https://doi.org/10.1111/mam.12173
Openshaw JJ, Hegde S, Sazzad HM, Khan SU, Hossain MJ, Epstein JH, Daszak P, Gurley ES, Luby SP (2017) Bat hunting and Bat–human interactions in Bangladeshi villages: implications for zoonotic disease transmission and Bat conservation. Transbound Emerg Dis 64(4):1287–1293. https://doi.org/10.1111/tbed.12505
Paddock RC, Sijabat DM (2020) Where Bats Are Still on the Menu, if No Longer the Best Seller. International New York Times. https://www.nytimes.com/2020/05/13/world/asia/coronavirus-bats-market-Indonesia.html
Pan American Health Organization (2024) Regional Information System for Epidemiological Surveillance of Rabies (SIRVERA). Available online: https://sirvera.panaftosa.org.br. (Accessed 15 May 2024)
Paterson BJ, Butler MT, Eastwood K, Cashman PM, Jones A, Durrheim DN (2014) Cross sectional survey of human-bat interaction in australia: public health implications. BMC Public Health 14:1–8. https://doi.org/10.1186/1471-2458-14-58
Paton NI, Leo YS, Zaki SR, Auchus AP, Lee KE, Ling AE, Chew SK, Ang B, Rollin PE, Umapathi T, Sng I (1999) Outbreak of Nipah-virus infection among abattoir workers in Singapore. Lancet 354(9186):1253–1256. https://doi.org/10.1016/S0140-6736(99)04379-2
Pavlovich SS, Darling T, Hume AJ, Davey RA, Feng F, Mühlberger E, Kepler TB (2020) Egyptian rousette IFN-ω subtypes elicit distinct antiviral effects and transcriptional responses in conspecific cells. Front Immunol 11:435. https://doi.org/10.3389/fimmu.2020.00435
Phillips AM, Gonzalez LO, Nekongo EE, Ponomarenko AI, McHugh SM, Butty VL, Levine SS, Lin YS, Mirny LA, Shoulders MD (2017) Host proteostasis modulates influenza evolution. Elife 6:e28652. https://doi.org/10.7554/eLife.28652
Pillai VS, Krishna G, Veettil MV (2020) Nipah virus: past outbreaks and future containment. Viruses 12(4):465. https://doi.org/10.3390/v12040465
Pimentel MF, Nassarden SM, Cândido SL, Dutra V, Nakazato L (2022) Genotyping of rabies positive samples isolated from animals in Mato Grosso and Rondônia–Brazil. Infect Genet Evol 103:105336. https://doi.org/10.1016/j.meegid.2022.105336
Platto S, Xue T, Carafoli E (2020) COVID19: an announced pandemic. Cell Death Dis 11(9):799. https://doi.org/10.1038/s41419-020-02995-9
Platto S, Wang Y, Zhou J, Carafoli E (2021) History of the COVID-19 pandemic: origin, explosion, worldwide spreading. Biochem Biophys Res Commun 538:14–23. https://doi.org/10.1016/j.bbrc.2020.10.087
Plowright RK, Peel AJ, Streicker DG, Gilbert AT, McCallum H, Wood J, Baker ML, Restif O (2016) Transmission or within-host dynamics driving pulses of zoonotic viruses in reservoir–host populations. PLoS Negl Trop Dis 10(8):e0004796. https://doi.org/10.1371/journal.pntd.0004796
Primack RB, Morrison RA (2013) Extinction, causes of, pp.401–412. https://doi.org/10.1016/B978-0-12-384719-5.00050-2
ProMED-mail (2014) Australian bat lyssavirus - Australia (03): (NS) flying fox, human exp, Archive Number: 20141025.2900858. ProMED-mail. (Accessed 25 May 2024). http://www.promedmail.org/
Quader MA, Agrawal S, Kervyn M (2017) Multi-decadal land cover evolution in the Sundarban, the largest mangrove forest in the world. Ocean & coastal management 139:113– 24. https://doi.org/10.1016/j.ocecoaman.2017.02.008
Ramírez-Fráncel L, García‐Herrera, Lv, Losada‐Prado S, Reinoso‐Flórez G, Sánchez‐Hernández A, Estrada‐Villegas S, Lim BK, Guevara G (2022) Bats and their vital ecosystem services: a global review. Integrative Zoology 17(1):2–3. https://doi.org/10.1111/1749-4877.12552
Rana S, Singh S (2015) Nipah virus: effects of urbanization and climate change. In3rd International Conference on biological, chemical & environmental sciences (BCES-2015) (pp. 64–68). https://iicbe.org/upload/7575C0915051.pdf
Reyes-del Valle J, Chávez-Salinas S, Medina F, Del Angel RM (2005) Heat shock protein 90 and heat shock protein 70 are components of dengue virus receptor complex in human cells. J Virol 79(8):4557–4567. https://doi.org/10.1128/jvi.79.8.4557-4567.2005
Richards DR, Friess DA (2016) Rates and drivers of mangrove deforestation in Southeast Asia, 2000–2012. Proceedings of the National Academy of Sciences 113(2):344-9. https://doi.org/10.1073/pnas.1510272113
Roberts BJ, Catterall CP, Eby P, Kanowski J (2012) Long-distance and frequent movements of the flying-fox Pteropus poliocephalus: implications for management. PLoS ONE 7(8):e42532. https://doi.org/10.1371/journal.pone.0042532
Russo D, Bosso L, Ancillotto L (2018) Novel perspectives on Bat insectivory highlight the value of this ecosystem service in farmland: research frontiers and management implications. Agric Ecosyst Environ 266:31–38. https://doi.org/10.1016/j.agee.2018.07.024
Salazar G, Mills M, Veríssimo D (2019) Qualitative impact evaluation of a social marketing campaign for conservation. Conserv Biol 33(3):634–644. https://doi.org/10.1111/cobi.13218
Salinas-Ramos VB, Mori E, Bosso L, Ancillotto L, Russo D (2021) Zoonotic risk: one more good reason why cats should be kept away from bats. Pathogens 10(3):304. https://doi.org/10.3390/pathogens10030304
Santos ND, Paz ES, Carneiro ID, Franke CR (2019) Evaluation of bat-related knowledge, perceptions, and practices in an urban community: A strategy for conservation biology and health promotion. Brazilian J Biol Sci 6(13):347–358. https://doi.org/10.21472/bjbs.061303
Schneeberger K, Voigt CC (2016) Zoonotic viruses and conservation of bats. Bats in the Anthropocene: Conservation of bats in a changing world pp.263– 92. https://doi.org/10.1007/978-3-319-25220-9_10
Schountz T, Baker ML, Butler J, Munster V (2017) Immunological control of viral infections in bats and the emergence of viruses highly pathogenic to humans. Front Immunol 8:1098. https://doi.org/10.3389/fimmu.2017.01098
Schulze MD, Seavy NE, Whitacre DF (2000) A comparison of the phyllostomid Bat assemblages in undisturbed Neotropical forest and in forest fragments of a Slash-and-Burn farming mosaic in petén, Guatemala 1. Biotropica 32(1):174–184. https://doi.org/10.1111/j.1744-7429.2000.tb00459.x
Seronello S, Montanez J, Presleigh K, Barlow M, Park SB, Choi J (2011) Ethanol and reactive species increase basal sequence heterogeneity of hepatitis C virus and produce variants with reduced susceptibility to antivirals. PLoS ONE 6(11):e27436. https://doi.org/10.1371/journal.pone.0027436
Sheldon BC, Verhulst S (1996) Ecological immunology: costly parasite defences and trade-offs in evolutionary ecology. Trends Ecol Evol 11(8):317–321. https://doi.org/10.1016/0169-5347(96)10039-2
Simmons NB, Cirranello AL (2022) Bat Species of the World: A taxonomic and geographic reference. https://batnames.org
Singh R, Kølvraa S, Bross P, Christensen K, Gregersen N, Tan Q, Jensen UB, Eiberg H, Rattan SI (2006) Heat-shock protein 70 genes and human longevity: a view from Denmark. Ann N Y Acad Sci 1067(1):301–308. https://doi.org/10.1196/annals.1354.040
Sohayati AR, Hassan L, Sharifah SH, Lazarus K, Zaini CM, Epstein JH, Naim NS, Field HE, Arshad SS, Aziz JA, Daszak P (2011) Evidence for Nipah virus recrudescence and serological patterns of captive Pteropus vampyrus. Epidemiol Infect 139(10):1570–1579. https://doi.org/10.1017/S0950268811000550
Srivastava P (2002) Roles of heat-shock proteins in innate and adaptive immunity. Nat Rev Immunol 2(3):185–194. https://doi.org/10.1038/nri749
Subudhi S, Rapin N, Dorville N, Hill JE, Town J, Willis CK, Bollinger TK, Misra V (2018) Isolation, characterization and prevalence of a novel gammaherpesvirus in Eptesicus fuscus, the North American big brown Bat. Virology 516:227–238. https://doi.org/10.1016/j.virol.2018.01.024
Subudhi S, Rapin N, Misra V (2019) Immune system modulation and viral persistence in bats: Understanding viral spillover. Viruses 11(2):192. https://doi.org/10.3390/v11020192
Tabish SA, Nabil S (2022) An age of emerging and reemerging pandemic threats. Health 14(10):1021–1037. https://doi.org/10.4236/health.2022.1410073
Tanalgo KC, Sritongchuay T, Agduma AR, Cruz KC, Hughes AC (2023) Are we hunting bats to extinction? Worldwide patterns of hunting risk in bats are driven by species ecology and regional economics. Biol Conserv 279:109944. https://doi.org/10.1016/j.biocon.2023.109944
Teeling EC, Springer MS, Madsen O, Bates P, O’brien SJ, Murphy WJ (2005) A molecular phylogeny for bats illuminates biogeography and the fossil record. Science 307(5709):580–584. https://doi.org/10.1126/science.1105113
Tian J, Sun J, Li D, Wang N, Wang L, Zhang C, Meng X, Ji X, Suchard MA, Zhang X, Lai A (2022) Emerging viruses: Cross-species transmission of coronaviruses, filoviruses, henipaviruses, and rotaviruses from bats. Cell Rep 39(11). https://doi.org/10.1016/j.celrep.2022.110969
Valitutto MT, Aung O, Tun KY, Vodzak ME, Zimmerman D, Yu JH, Win YT, Maw MT, Thein WZ, Win HH, Dhanota J (2020) Detection of novel coronaviruses in bats in Myanmar. PLoS ONE 15(4):e0230802. https://doi.org/10.1371/journal.pone.0230802
Veettil BK, Quang NX (2019) Mangrove forests of cambodia: recent changes and future threats. Ocean Coastal Manage 181:104895. https://doi.org/10.1016/j.ocecoaman.2019.104895
Vora NM, Osinubi MO, Davis L, Abdurrahman M, Adedire EB, Akpan H, Aman-Oloniyo AF, Audu SW, Blau D, Dankoli RS, Ehimiyein AM (2020) Bat and lyssavirus exposure among humans in area that celebrates Bat festival, nigeria, 2010 and 2013. Emerg Infect Dis 26(7):1399. https://doi.org/10.3201/eid2607.191016
Weber N, Nagy M, Markotter W, Schaer J, Puechmaille SJ, Sutton J, Dávalos LM, Dusabe MC, Ejotre I, Fenton MB, Knörnschild M (2023) Robust evidence for bats as reservoir hosts is lacking in most African virus studies: a review and call to optimize sampling and conserve bats. Biol Lett 19(11):20230358. https://doi.org/10.1098/rsbl.2023.0358
Weinberg M, Yovel Y (2022) Revising the paradigm: are bats really pathogen reservoirs or do they possess an efficient immune system? Iscience 25(8)
Wilkinson GS, South JM (2002) Life history, ecology and longevity in bats. Aging Cell 1(2):124–131. https://doi.org/10.1046/j.1474-9728.2002.00020.x
Wirsiy FS, Boock AU, Akoachere JF (2021) Assessing the determinants of Ebola virus disease transmission in Baka community of the tropical rainforest of Cameroon. BMC Infect Dis 21:1–1. https://doi.org/10.1186/s12879-021-06011-z
World Health Organization (2023a) El Niño Southern Oscillation (ENSO), World Health Organization website (Accessed 2 May 2024). https://www.who.int/news-room/fact-sheets/detail/el-nino-southern-oscillation-(enso)
World Health Organization (2023b) Disease Outbreak News; Nipah Virus Infection in India, World Health Organization website (Accessed 6 May 2024). https://www.who.int/emergencies/disease-outbreak-news/item/2023-DON490
World Health Organization (2022) WHO to identify pathogens that could cause future outbreaks and pandemics, World Health Organization website (accessed 28 May 2024). https://www.who.int/news/item/21-11-2022-who-to-identify-pathogens-that-could-cause-future-outbreaks-and-pandemics#:~:text=The%20current%20list%20includes%20COVID,fever%2C%20Zika%20and%20Disease%20X
Xiao X, Newman C, Buesching CD, Macdonald DW, Zhou ZM (2021) Animal sales from Wuhan wet markets immediately prior to the COVID-19 pandemic. Sci Rep 11(1):11898
Xie J, Li Y, Shen X, Goh G, Zhu Y, Cui J, Wang LF, Shi ZL, Zhou P (2018) Dampened STING-dependent interferon activation in bats. Cell Host Microbe 23(3):297–301. https://doi.org/10.1016/j.chom.2018.01.006
Ye ZW, Yuan S, Yuen KS, Fung SY, Chan CP, Jin DY (2020) Zoonotic origins of human coronaviruses. Int J Biol Sci 16(10):1686. https://doi.org/10.7150/ijbs.45472
Yuen KY, Fraser NS, Henning J, Halpin K, Gibson JS, Betzien L, Stewart AJ (2021) Hendra virus: epidemiology dynamics in relation to climate change, diagnostic tests and control measures. One Health 12:100207. https://doi.org/10.1016/j.onehlt.2020.100207
Zaki AM, Van Boheemen S, Bestebroer TM, Osterhaus AD, Fouchier RA (2012) Isolation of a novel coronavirus from a man with pneumonia in Saudi Arabia. N Engl J Med 367(19):1814–1820. https://doi.org/10.1056/NEJMoa1211721
Zhang YZ, Holmes EC (2020) A genomic perspective on the origin and emergence of SARS-CoV-2. Cell 181(2):223–227. https://doi.org/10.1016/j.cell.2020.03.035
Zhang G, Cowled C, Shi Z, Huang Z, Bishop-Lilly KA, Fang X, Wynne JW, Xiong Z, Baker ML, Zhao W, Tachedjian M (2013) Comparative analysis of Bat genomes provides insight into the evolution of flight and immunity. Science 339(6118):456–460. https://doi.org/10.1126/science.1230835
Zheng J (2020) SARS-CoV-2: an emerging coronavirus that causes a global threat. Int J + + Biol Sci 16(10):1678. https://doi.org/10.7150/ijbs.45053
Zhou P, Cowled C, Mansell A, Monaghan P, Green D, Wu L, Shi Z, Wang LF, Baker ML (2014) IRF7 in the Australian black flying fox, Pteropus alecto: evidence for a unique expression pattern and functional conservation. PLoS ONE 9(8):e103875. https://doi.org/10.1371/journal.pone.0103875
Zhu N, Zhang D, Wang W, Li X, Yang B, Song J, Zhao X, Huang B, Shi W, Lu R, Niu P (2020) A novel coronavirus from patients with pneumonia in china, 2019. N Engl J Med 382(8):727–733. https://doi.org/10.1056/NEJMoa2001017
Acknowledgements
The authors recognize the use of online software, Biorender and PRISMA, for making figures and the generation of flow diagrams for systematic review, respectively.
Funding
No funding has been received for this work.
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article
Cite this article
Mahanta, K., Sulabh, S. Zoonotic spillovers: How climate change, habitat destruction, and bushmeat trade might amplify bat-driven viral disease risks. Eur J Wildl Res 71, 72 (2025). https://doi.org/10.1007/s10344-025-01951-2
Received:
Revised:
Accepted:
Published:
Version of record:
DOI: https://doi.org/10.1007/s10344-025-01951-2
Share this article
Anyone you share the following link with will be able to read this content:
Sorry, a shareable link is not currently available for this article.
Provided by the Springer Nature SharedIt content-sharing initiative
Keywords
Profiles
- Sourabh Sulabh View author profile